背景及概述[1]
促肾上腺皮质激素简称促皮质素(adrenocorticotropichormone,adrenocorticotropin,ACTH)是从哺乳动物脑垂体前叶中提取到的一个多肽激素。本品也可用合成法制造。促皮质素为白色或淡黄色冻干制剂。易溶于水、乙醇。在弱酸中较稳定,强酸或强碱中容易失活。猪促皮质素由39个氨基酸组成,其结构式见上。牛、羊、猪及人促皮质素的结构相差不多,人促皮质素第31位为“丝氨酸”,猪为“亮氨酸”,在25位的“天冬氨酸”和33位的“谷氨酸”,有时变为“天冬酰胺”及“谷氨酰胺”。促皮质素的活性与近N-端的肽链结构有密切关系。其作用机理是作用于靶细胞膜上的受体。促皮质素能维持肾上腺皮质的正常功能,促进皮质激素的合成和分泌。但当肾上腺皮质本身受损,分泌功能发生严重障碍时,ACTH就不能奏效,所以本品也可作为诊断垂体和肾上腺皮质功能的药物。临床上主要用于胶原病,例如风湿性关节炎、红斑狼疮、干癣,也用于过敏症例如严重喘息、药物过敏、荨麻症等。近年发现ACTH也存在于中枢神经系统,与人的记忆和行为有联系,可以改善老年人以及智力迟钝儿童的学习和记忆能力。促皮质素的制剂类型甚多。有长效的锌促皮质素,明胶促皮质素。羧甲基纤维促皮质素。此外尚有合成的长效衍生物丝-赖促皮质素(18肽)、甘-精促皮质肽(18肽)和人促皮质素结构相同的25肽等。锌促皮质素肌注后6小时见效,12到30小时作用达最高峰。持续时间达48小时。长期使用ACTH,副作用有浮肿、肌肉衰弱、神经过敏、血压升高、高血糖,偶有过敏休克。在用药期间应少进食盐,并进行血中钾、钠离子的检查,尿中17-酮体检查。在终止用药时应采取逐步减少剂量的方法,以防止肾上腺功能减退。有精神病、高血压、糖尿、肾炎以及孕妇等均忌用。剂量:初次剂量为每天40~80u,以后逐步减到20u。。
结构[2]
ACTH是一种线性多肽,与α-促黑色素细胞激素(α-melanocytestimulatinghormone,α-MSH),促皮质激素样中叶肽等均来源于前阿黑皮素前体。
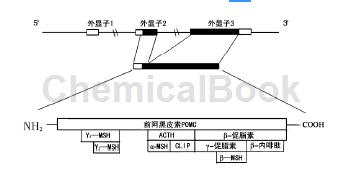
ACTH含39个氨基酸,其中1~24位氨基酸是构成生物活性的必备氨基酸,25~39位为保护性氨基酸。其中ACTH的20到24残基对其生物利用度有负调节作用,并且对促皮质激素样中叶肽的作用有重要意义,而促皮质激素样中叶肽可以抑制N-甲基-D-天门冬氨酸(NMDA受体),并能强化突触后GABA受体和AMPA受体的作用[5]。ACTH主要产自垂体前叶,垂体中叶中的ACTH主要作为生物合成的中间产物存在。垂体来源的ACTH与血清ACTH浓度关系密切,而大脑中的ACTH则主要来源于下丘脑的旁分泌。人体内ACTH广泛存在于垂体之外的诸多组织,如免疫细胞(扁桃体T细胞、外周血单核细胞)、胎盘滋养母细胞等。在中枢神经系统内ACTH的分布也很广泛。ACTH在全身各大系统的广泛分布提示ACTH作用的广泛性。
应用[2-3]
1.ACTH与黑皮质素受体
ACTH受体来源黑皮质素受体(melanocortinreceptor,MCR)家族。MCR家族是一组G蛋白偶联受体,共有5个成员,其中只有MC2R是ACTH的特异性受体,但ACTH是所有MCR家族的内源性激动剂。MC2R几乎仅存于肾上腺,但与MC2R作用相关的黑皮质素受体辅助蛋白与黑皮质素受体辅助蛋白-2的分布比MC2R更为广泛,黑皮质素受体辅助蛋白-2在大脑亦有分布。MC2R与黑皮质素受体辅助蛋白形成复合物则在束状带浓度最高,主要作用是促进皮质激素生成;MC2R与黑皮质素受体辅助蛋白-2形成复合物则在全肾上腺均呈低浓度分布,作用主要与肾上腺细胞的分化相关。黑皮质素受体辅助蛋白具有下调除MC2R以外的其他4个黑皮质素受体家族成员的作用,并且可以调节黑皮质素受体的表达及信号传导能力,由此猜测黑皮质素受体辅助蛋白在调节ACTH与MCR受体家族其他成员的相互作用中发挥一定作用。MC5R主要分布于淋巴细胞和外分泌腺,而MC1RMC3R、MC4R在大脑分布则更具优势。在中枢系统,MC1R多分布在脑水管周灰质、星形细胞、施旺细胞及微血管内皮细胞上。MC3R分布在下丘脑及边缘系统。MC4R分布更广,在皮质、丘脑、下丘脑、脑干、脊髓及星形细胞上均有分布。ACTH与α-MSH和这些受体的结合位点并不完全一致,推测ACTH对神经系统有区别于α-MSH的作用。
2.ACTH与脑的发育
1)脑发育过程中ACTH的表达ACTH在中枢系统的表达较早,前阿黑皮素/ACTH在10.5d胎龄大鼠的背侧间脑即有表达,12.5d胎龄大鼠的下丘脑亦可检测到前阿黑皮素/ACTH的存在,随即在第13.5~15.5d胎龄大鼠的间脑、中脑、后脑、末脑等迅速形成大量致密神经纤维束。对这段时期出现的神经纤维形成,猜测可能与ACTH的作用相关。通过原位杂交的办法对14d胎龄到生后27d的evans大鼠神经中枢尤其是脑部的MC3R和MC4RmRNA进行检测和定位,发现在第14d胎龄即可检测到MC4R,在胎鼠大脑以MC4R为主,在乳鼠大脑以MC3R为主。MC4R在交感神经节、躯体感觉和内脏感觉神经核团、一些颅神经节(如三叉神经节、膝状神经节等)、中脑、间脑、小脑、脑干、大脑皮层、脊髓等不同区域的不同发育时期均有所分布;MC3R虽然在中枢系统出现较晚,同样分布广泛。
2)胚胎期ACTH的来源与调节作用母体源性的ACTH几乎无法通过胎盘,因此在胚胎发育早期,胎盘是胎儿ACTH最重要的来源之一。白细胞抑制因子(leucocyteinhibitoryfactor,LIF)、白介素-6、制瘤素M、CRH均可诱导ACTH分泌的增加,其中LIF对ACTH的诱导作用最强。母鼠体内的LIF水平在孕14.5d时升高,母源性LIF与胎盘微绒毛上滋养母细胞母体侧白细胞抑制因子受体及gp130蛋白结合,激活JAK/STAT3信号传导通路,促使胎盘滋养母细胞表达大量合成分泌ACTH。在胚胎期ACTH对神经系统发育的作用是多途径的,ACTH可与胎鼠有核红细胞上的黑皮质素受体MC2R/MC5R结合,促进胎鼠有核红细胞产生大量LIF。胚胎期LIF对神经元发生,神经干细胞、祖细胞增殖分化及星型胶质细胞的分化具有促进作用,可通过gp130信号通路使脑室带的祖细胞迅速进入新的细胞分裂周期而无需缩短细胞周期的时间,进而促进神经元发生和分化进程。而ACTH对胚胎期LIF的水平有调节作用。胎鼠有核红细胞对ACTH的敏感性在孕15.5d时达到高峰,随后降低,使胎鼠在15.5d胎龄时血清和脑脊液中的LIF水平达到一个高峰,在这一时期胎鼠大脑新皮质的脑室带细胞数目随之增加。ACTH通过从母体到胎体的LIF-ACTH-LIF信号转导,使胎鼠免于受母体LIF水平大幅波动的影响,保证大脑发育处于一个较为稳定有利的环境。
3)ACTH与HPAA的其他成员作为HPAA系统的重要成员,ACTH对HPA轴的调节作用是ACTH所具备的作用中最重要的成分。在妊娠中晚期,HPA轴逐渐形成并具有稳定活性,ACTH、CRH和糖皮质激素的相互调节,使彼此处于一个较为稳定的水平,共同发挥其对大脑发育的作用、维持发育后大脑功的稳定以及调节应激性反应对脑功能影响的作用。近年来的许多研究均认为发生在HPA轴形成的敏感时期内的环境因素及糖皮质激素、CRH水平变化均会对一生的健康产生深远的影响。因HPA轴对应激反应及损伤的修复有重要作用,HPA系统的“调节反馈程序”在发育过程中发生的“程序”变化可能导致HPA轴对应激性损伤的调节反应发生改变从而影响糖皮质激素、CRH水平变化及机体的抗损伤及修复能力。ACTH对脑发育的影响很大部分是通过调节CRH和糖皮质激素水平而实现的,而CRH和糖皮质激素对大脑的发育也有各自特殊的作用。用cRNA探针对14d胎龄至生后16d大鼠脑糖皮质激素受体mRNA进行检测时发现,早在14d胎龄时(即海马中隔形成的时期)就可检测到糖皮质激素受体mRNA的存在,到16d胎龄时下丘脑室旁核的糖皮质激素受体表达就已经很明显了,而此时下丘脑室旁核中CRH基因的表达尚未出现,这也许提示糖皮质激素受体对神经元分化有一定影响。在胚胎晚期,糖皮质激素受体已经广泛表达于丘脑、海马、杏仁核和周质区。在新生乳鼠的下丘脑室旁核、海马趾的CA1/CA2区、梨状皮质和背侧梨状内核、杏仁核、视交叉上核也均存在丰富的糖皮质激素受体。此外,切除孕期母狗双侧肾上腺或幼狗双侧肾上腺后,幼狗脑重量和髓鞘形成均增加,尤其是前脑和后脑。并且这种改变在给与生理需要量的糖皮质激素后即可反转。此结果暗示糖皮质激素在脑发育尤其前脑边缘系统的发育中具促进作用。在另一系列研究中,胚胎发育早期实行母体双侧肾上腺切除术可以导致海马齿状回和CA1亚区细胞数量的减少以及海马发育的迟滞。这一结果同样支持了糖皮质激素在神经系统中存在促进神经元发生的作用。但是过高水平的糖皮质激素通过激活一氧化氮途径可产生过量的活性氧、升高细胞内钙浓度,使神经元过度极化,从而导致神经元毒性;高水平的糖皮质激素亦可使神经产生退行性变从而对中枢神经系统产生危害。在胚胎发育期间ACTH出现较早,在早期即可发挥其对糖皮质激素水平的调节作用,使其处于一个较为稳定合理的水平,与糖皮质激素共同促进神经系统的发育,并有效抑制糖皮质激素水平的过度升高,有效地避免过高激素水平对发育中的大脑造成损伤。
3.ACTH与神经递质的相互作用
ACTH对神经内分泌的调节涉及多种神经递质的共同参与,这些神经递质包含了兴奋性神经递质及抑制性神经递质。试验中观察到弓状核中前阿黑皮素阳性神经元可以释放兴奋性氨基酸谷氨酸Glu或者抑制性氨基酸GABA。兴奋性神经递质和抑制性神经递质均可对HPAA的活性产生调节。Glu、红藻氨酸、使君子氨酸等兴奋性氨基酸或其受体的激动剂可直接作用于下丘脑室旁核而促ACTH的合成释放;封闭Glu的受体,如NMDA受体、α-氨基羟甲基恶唑丙酸AMPA受体、亲代谢型受体等,均可以观察到ACTH的释放受到抑制。兴奋性氨基酸与惊厥、癫痫的发生相关,在临床治疗中ACTH表现出良好的抗惊厥作用,这提示ACTH与兴奋性氨基酸作用的失平衡参与了中枢神经系统功能紊乱性疾病的发生。另有试验对6个月的大鼠进行颅内注射NMDA,观察到生后立即使用Org2766[ACTH(4-9)类似物]的大鼠胆碱能神经元受损程度有所减轻,神经元及神经胶质细胞对兴奋毒性造成的损伤的抵制能力也有所增强。此试验不仅说明了ACTH对胆碱能神经元损伤修复作用与其对兴奋性氨基酸的抑制性调节作用有关,并且提示在发育早期ACTH可以促进神经内在保护机制的发育和成熟,从而长远地影响神经元的兴奋可塑性。ACTH对中枢神经系统的内在保护不仅得益于ACTH与兴奋性神经递质的作用,同时也得益于ACTH与抑制性神经递质的相互作用。近年来研究发现,ACTH的抗惊厥作用与GABA受体阻断有密切关系[28]。与兴奋性氨基酸Glu相反,抑制性氨基酸GABA不仅直接引发神经元的抑制性突触后电位,还可通过直接作用于下丘脑室旁核的CRH神经元来抑制HPA轴活性。在下丘脑水平,GABA与ACTH的相互作用十分复杂。在清醒大鼠的内侧视前区注射GABA受体激动剂蝇蕈醇,可以观察到ACTH水平的升高和下丘脑室旁核中c-Fox表达的增加。在背内侧下丘脑进行蝇蕈醇预处理,ACTH水平升高不明显;注射GABA受体拮抗剂,则ACTH水平有显著升高。可见在丘脑的不同区域GABA与ACTH的相互作用不尽相同。
4.ACTH与婴儿痉挛
婴儿痉挛是一种与年龄相关同时伴有智力运动发育倒退现象的癫痫综合征。长期以来,ACTH作为婴儿痉挛治疗的一线药物,其作用机制并不清楚。目前有人提出婴儿痉挛的发病机制与CRH水平升高有关:发生在胚胎发育早期的应激性事件可引起糖皮质激素受体基因甲基化,导致HPA轴负反馈调节能力下降,HPA系统的“调节反馈程序”异常,从而引起CRH的过度表达,并最终导致婴儿痉挛的发生。而ACTH可以通过HPA轴的负反馈作用(ACTH本身的负反馈作用和ACTH引起皮质醇升高后皮质醇的负反馈作用)以及ACTH对杏仁核神经元等的直接作用下调CRH的表达,从而降低CRH水平,防止惊厥发生。这似乎很好地解释了为何婴儿痉挛对传统的抗癫痫药物并不敏感,但这无法完全解释ACTH和糖皮质激素对婴儿痉挛良好的治疗效果及为何部分婴儿痉挛患儿对ACTH和激素治疗并不敏感。对MC4R启动子的单核苷酸多态性进行进一步研究,发现MC4R启动子等位基因rs11872992的多态性与婴儿痉挛患者对ACTH治疗的敏感程度相关,这一结果暗示ACTH对婴儿痉挛的治疗与ACTH和脑MC4R的作用相关。另有研究发现MC2R基因启动子的多态性影响了ACTH对婴儿痉挛治疗的有效性,ACTH对携带TCCT基因型的婴儿痉挛患儿有更好的治疗效果,进一步的体外功能学研究证实MC2R基因启动子TCCT单体型可明显提高MC2R启动子的转录效率(TCCT型启动子转录效率是TCCC型的4倍)。这些均证实了ACTH与MCR的互作用不仅仅只是促进了MSH与MCR的作用,其与MCR还可通过其他多种神经内分泌途径发挥对神经系统的影响。在对婴儿痉挛的治疗中,一方面,ACTH表现出良好的抗惊厥效果;另一方面,ACTH对病后的智力运动发育倒退亦有所改善。这些均提示,在对婴儿痉挛的治疗中ACTH的影响是多途径的,涉及其中的机制也许不仅仅是ACTH对神经递质和神经营养因子的调节作用,或其过与MCR家族成员的相互作用影响HPAA其他成员(CRH、糖皮质激素)的水平而发挥作用。近年来的多项研究还发现,ACTH的抗惊厥作用与其促进神经甾体的分泌合成有关,ACTH促进皮质酮的分泌,其中去氧皮质酮(盐皮质激素的一种)可转化为5α-3α-四氢脱氧皮质酮(GABAA受体的别构调节剂),通过阻断GABAA受体的作用而发挥抗惊厥作用。这一发现也支持了ACTH是一个作用广泛的神经调节因子。
主要参考资料
[1] 中国医学百科全书·十八药物学与药理学
[2] 促肾上腺皮质激素与中枢神经系统发育研究进展
[3] 促肾上腺皮质激素治疗肾性蛋白尿的研究