背景[1][2][3][4]
GSK-3β抗体(兔单抗)是用GSK-3β对兔进行系统免疫然后筛选出兔GSK-3β致敏B细胞后与骨髓肿瘤细胞杂交最后制得既能产生单一GSK-3β抗体,又能无限增殖的GSK-3β抗体杂种细胞系。糖原合成酶激酶3也称为GSK3B,是一种酶,在人中由编码GSK3B基因。在小鼠中,酶由GSK-3β基因编码。GSK3β的异常调节和表达与双相情感障碍的易感性增加有关。糖原合成酶激酶-3(GSK-3)是一种脯氨酸定向的丝氨酸-苏氨酸激酶,最初被鉴定为糖原合成酶的磷酸化和灭活剂。
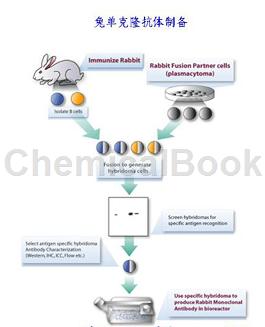
两种同种型α(GSK3A)和β显示出高度的氨基酸同源性。GSK3B参与能量代谢,神经细胞发育和体型形成。它可能是缺血性中风的新治疗靶点。小鼠中GSK-3β基因座的纯合破坏导致妊娠中期的胚胎致死率。这种致死性表型可以通过抑制肿瘤坏死因子来挽救。ERK1/2的药理学抑制恢复了结节性硬化模型中的GSK3β活性和蛋白质合成水平。
研究应用[5][6]
1.用于甘草黄酮对2型糖尿病大鼠肝脏中GSK-3β蛋白表达的影响
研究甘草黄酮(LF)对2型糖尿病大鼠肝脏中GSK-3β蛋白表达的影响。采用高脂高糖饲料喂养结合一次性小剂量腹腔注射链脲佐菌素(STZ)的方法建立2型糖尿病(T2DM)实验大鼠模型。将实验大鼠分为正常对照组(CON)、糖尿病模型组(DM)、甘草黄酮治疗组(LF)和阳性对照组(PC)。分别以溶剂(1,2-丙二醇)、甘草黄酮[300 mg/(kg·d)]或吡格列酮[10 mg/(kg·d)]连续灌胃6周。
采用蛋白免疫印迹(Westernblotting)方法检测各组大鼠肝脏中GSK-3β的蛋白表达量。研究结果显示,DM组大鼠肝脏组织中GSK-3β的蛋白表达量较CON组显著升高(P<0.01);其在LF组和PC组的表达量与DM组相比均显著降低(P<0.05)。上述结果表明,LF能显著降低2型糖尿病大鼠肝脏中GSK-3β的蛋白表达量,这可能是其改善胰岛素抵抗的分子机制之一。
2.用于GSK-3β/β-catenin信号通路在磨损颗粒诱导假体周围骨溶解中的作用及机制研究
方法:雌性C57BL/6小鼠84只,采用随机数字表法分为4组:对照组(Control),Ti颗粒组(Ti组),Li Cl低剂量治疗组(L-Li Cl组)和高剂量治疗组(H-Li Cl组),每组21只小鼠。Control组仅接受假手术,术后每天灌胃300μl等渗盐水;Ti组在小鼠颅骨表面植入Ti颗粒,术后每天灌胃300μl等渗盐水;L-Li Cl和H-Li Cl组,Ti颗粒植入后,分别灌胃50或200 mg kg-1d-1Li Cl。给药2周后,使用CO2箱处死小鼠,保留颅骨及外周血,分别行micro-CT、组织病理学染色、RT-PCR、酶联免疫吸附试验(ELISA)、放射免疫分析、血常规及血生化等检测。
结果:Micro-CT结果显示治疗组颅骨骨溶解现象明显减轻,骨密度、骨体积、骨体积分数L-Li Cl组和H-Li Cl组分别比Ti组增加23.8%、51.7%,18.1%、45.8%,21.7%、43.9%;骨陷窝数减少了37.5%和59.8%,差异有统计学意义(p<0.05)。H&E染色结果表明,Ti组骨膜内有大量巨噬样细胞,L-Li Cl组和H-Li Cl组炎症反应明显减轻。Ti组骨溶解面积(BES)和颅骨厚度(BT)分别为(0.13±0.03)mm2、(0.12±0.04)mm,L-Li Cl组和H-Li Cl组分别为(0.07±0.01)mm2、(0.04±0.008)mm2,(0.16±0.03)mm、(0.22±0.11)mm,与Ti组比较,差异有统计学意义(p<0.05)。抗酒石酸酸性磷酸酶染色(TRAP)结果显示,Ti组颅骨溶解侧可见大量TRAP阳性细胞,治疗后TRAP阳性细胞数量显著减少。
免疫组织化学染色结果表明Li Cl能增加p Ser9-GSK-3β、β-catenin、ALP、Osterix和OPG的表达,抑制RANKL、TNF-α、IL-1β和IL-6的表达,而不影响GSK-3β的表达。RT-PCR结果显示,Ti颗粒显著上调破骨细胞活化相关基因Cath-K、CTR、TRAP、OSCAR m RNA的表达,经治疗后,上述基因m RNA表达水平降低,与Ti组比较差异有统计学意义(p<0.05)。Ti颗粒能抑制β-catenin和axin-2基因m RNA的表达,阻断GSK-3β后β-catenin和axin-2基因m RNA的表达明显增加。
结论:氯化锂通过调节GSK-3β/β-catenin信号通路,促进新骨形成,抑制骨吸收,减轻磨损颗粒引起的炎症性反应,抑制骨溶解。第二部分钛颗粒调控GSK-3β/β-catenin信号通路抑制成骨细胞分化目的:观察Ti颗粒对成骨细胞分化、炎症因子释放以及RANKL和OPG表达的影响,探讨GSK-3β/β-catenin在磨损颗粒调控成骨细胞功能中的作用。
参考文献
[1] Stambolic V,Woodgett JR(Nov 1994)."Mitogen inactivation of glycogen synthase kinase-3 beta in intact cells via serine 9 phosphorylation".The Biochemical Journal.303(Pt 3):701–4.
[2] Lau KF,Miller CC,Anderton BH,Shaw PC(Sep 1999)."Molecular cloning and characterization of the human glycogen synthase kinase-3beta promoter".Genomics.60(2):121–8.
[3] Hoeflich KP,Luo J,Rubie EA,Tsao MS,Jin O,Woodgett JR(2000)."Requirement for glycogen synthase kinase-3beta in cell survival and NF-kappaB activation".Nature.406(6791):86–90.
[4] Luykx JJ,Boks MP,Terwindt AP,Bakker S,Kahn RS,Ophoff RA(Jun 2010)."The involvement of GSK3beta in bipolar disorder:integrating evidence from multiple types of genetic studies".European Neuropsychopharmacology.20(6):357–68.
[5] 甘草黄酮对2型糖尿病大鼠肝脏中GSK-3β蛋白表达的影响[J].金鑫,赵海燕,马永平.天然产物研究与开发.2014(03)
[6] GSK-3β/β-catenin信号通路在磨损颗粒诱导假体周围骨溶解中的作用及机制研究[D].耿德春.苏州大学.2016